

Ключевые выводы
Все биологические изменения, которые мы наблюдаем в природе, сводятся либо к сохранению, либо к утрате существующей генетической информации. Новые функциональные информационные блоки не возникают. Поэтому реальные процессы не соответствуют тому, что обычно подразумевается под термином «эволюция». Исходя из логики, практичности и стратегической ясности, в статье предлагается три шага:
- Отказаться от использования термина «микроэволюция», так как он вводит в заблуждение.
- Пересмотреть наше понимание изменчивости внутри «библейского рода» (барамина), сместив фокус с морфологии на генетическую информацию.
- Не пытаться жестко привязывать библейские роды к современным таксономическим категориям (вид, семейство, род), кроме как на уровне аксиоматических определений.
В популярных изложениях эволюционной теории часто звучит мысль: если небольшие изменения, наблюдаемые сегодня, экстраполировать на огромные промежутки времени, то можно объяснить превращение одноклеточного организма в человека. Креационистов же иногда упрекают в том, что они проводят условную «линию Мажино», за которую биологическая изменчивость якобы не может выйти. Критики говорят, что креационисты просто не допускают, чтобы комары превратились в слонов, что является чрезмерным упрощением. Сторонник неодарвинизма может возразить, что таких масштабных изменений и не ожидается увидеть в течение жизни одного человека. А представитель более современных течений может заявить, что наш временной срез слишком мал, чтобы зафиксировать появление принципиально новой формы.
Еще одна причина, по которой позиция креационизма иногда кажется уязвимой, — это впечатление, будто мы признаем изменчивость лишь потому, что вынуждены это делать перед лицом фактов, но при этом избегаем обсуждать ее возможные долгосрочные последствия, утверждая, что она «не заходит достаточно далеко». Со стороны может показаться, что мы постоянно передвигаем нашу «линию обороны» в зависимости от новых данных. Однако далее в статье будет показано, что такой взгляд ошибочен и является карикатурой. Настоящие пределы изменчивости определяются не произвольными границами, а фундаментальным уровнем генетической информации.
Понятие сотворенного рода (барамина)
Очевидно, что наблюдаемая изменчивость имеет свои границы. Возникает соблазн использовать этот факт как прямое доказательство существования изначально сотворенных родов, внутри которых и происходит вся вариация.
Однако такая аргументация может быть уязвима для критики, так как содержит элементы кругового рассуждения. Если креационист по определению считает всю изменчивость ограниченной «пределами рода», то как с помощью наблюдений доказать, что эти пределы реальны? Всякий раз, когда две разновидности происходят от общего предка, их автоматически относят к одному роду. Неудивительно, что эволюционисты требуют четкого, проверяемого определения «сотворенного рода». Без этого утверждение об ограниченной изменчивости рискует стать тавтологией. Стоит отметить, что круговое обоснование само по себе не дискредитирует концепцию. Например, определение естественного отбора («выживают наиболее приспособленные, а наиболее приспособленные — это те, кто выжил») также содержит логический круг, но при этом остается рабочей и проверяемой концепцией. Мы лишь хотим сказать, что круговые аргументы не могут служить независимым доказательством существования бараминов.
Возможно, такое независимое доказательство в принципе недостижимо по самой природе вещей, и это не является оправданием. Допустим, мы нашли останки острова после мощного взрыва — камни, песок, обломки деревьев. Восстановить их первоначальное взаимное расположение может быть принципиально невозможно. Однако это не мешает нам с высокой долей уверенности заключить, что наблюдаемая картина согласуется со свидетельством очевидца о взрыве, а не с каким-либо иным процессом.
Аналогично, мы можем показать, что наблюдаемые в природе явления хорошо согласуются с библейской концепцией изначально сотворенных родов и плохо согласуются с идеей универсальной эволюции. Наиболее эффективно это сделать, если обратить внимание не на степень морфологических изменений, а на их информационную и генетическую основу.
Суть проблемы — в качестве изменений, а не в их количестве. Дело не в том, что «поезду эволюции» не хватило времени доехать, а в том, что он изначально поехал не в ту сторону. Пределы изменчивости — наблюдаемые или нет — неизбежно проявят себя, когда в генофонде будет исчерпан запас функционально эффективной (телеономической) генетической информации. Понимание этого снимает с креациониста нелепый образ человека, который в панике отступает и перерисовывает границы при каждом новом открытии.
Этот же факт делает излишним и искусственное разделение на «микро-» и «макроэволюцию». Креационистам лучше полностью отказаться от этих терминов, которые только сеют путаницу. Слово «эволюция» обычно подразумевает изменения, которые в итоге превращают простейшее в человека или рептилию в птицу. В рамках такого понимания мы не видим эволюции вообще. Говоря «я принимаю микро-, но не макроэволюцию», мы невольно поддерживаем идею, что разница между ними — лишь в количестве и времени, что в корне неверно. Речь идет о разных типах изменений.
Это не просто спор о словах, а вопрос, имеющий психологическое и тактическое значение. Конечно, можно дать термин «микроэволюция» какое-то частное определение, но его обыденное значение и коннотации таковы, что уступка здесь неразумна. Микроэволюция как изменение, пусть малое, но ведущее к увеличению функциональной информации, в природе не наблюдалась.
Любопытно, что многие ведущие биологи теперь сами признают, что «макроэволюция» — это не просто «микроэволюция», растянутая во времени. В ноябре 1980 года в Чикагском музее естественной истории прошла конференция по макроэволюции с участием видных эволюционистов. Роджер Левин, освещая ее в журнале Science, написал:
- «Центральным вопросом конференции было: можно ли экстраполировать механизмы микроэволюции для объяснения макроэволюции? Рискуя упростить позиции некоторых участников, ответ — определенно нет.»[1]
Со ссылкой на генетика Франсиско Айалы было упомянуто:
- «…судя по современным высказываниям палеонтологов, я уверен, что мелкие изменения не накапливаются.»[2]
Учитывая, что данная статья по сути приходит к схожему выводу, ее едва ли можно назвать радикальной. Тем не менее, многие образованные люди по-прежнему верны формуле: «(Большое изменение) = (Малые изменения) × (Миллионы лет)».
Информационная природа жизни
Буквы на этой странице состоят из чернил и бумаги, подчиняющихся законам физики и химии, но сами законы не определяют информацию, которую эти буквы несут. Информация зависит от материального носителя для хранения и передачи, но не является его неотъемлемым свойством. Идеи этой статьи сначала возникли в сознании, а затем были перенесены на бумагу. Биологические молекулы также несут колоссальные объемы информации, которая не является их химическим свойством. Информация возникает из специфического порядка, из способа сборки «букв» генетического алфавита. Этот порядок должен быть задан извне и не вытекает из свойств самих молекул. Живые организмы передают эту информацию из поколения в поколение. Последовательность оснований в ДНК — это детальный план, определяющий свойства организма. Наследственные изменения являются прямым следствием изменений в этой информации. Гены можно рассматривать как «предложения», записанные на языке ДНК.
Теперь представим первую популяцию организмов на «первобытной Земле» эволюциониста. Эта гипотетическая клетка содержала бы некоторое количество генетической информации, но неизмеримо меньшее, чем в генофонде ее предполагаемых потомков — например, людей. Эволюционист должен предположить, что эта короткая «телеграмма» стала родоначальницей целых «энциклопедий» новых осмысленных и полезных генетических «предложений». Единственным источником таких новых предложений в конечном счете могли быть только мутации.[3]
Возвращаясь к аналогии с печатной страницей, генетическая информация копируется при размножении, подобно тому как пишущая машинка воспроизводит текст. Мутация — это случайная ошибка копирования. Хотя большинство таких ошибок вредны или бессмысленны, эволюционисты полагают, что в определенных условиях среды они могут оказаться полезными, давая организму преимущество в выживании. Далее мы рассмотрим информационную основу других механизмов изменчивости и увидим, почему они не порождают новых «предложений», вынуждая эволюционистов апеллировать к мутациям.
1. Менделевская (комбинативная) изменчивость
Этот механизм отвечает за большинство новых разновидностей, получаемых в селекции и наблюдаемых в природе. Половое размножение позволяет по-разному комбинировать уже существующие пакеты генетической информации, но не создает новых. Например, все породы собак были выведены из «дикого типа» путем отбора и изоляции желаемых признаков в разных линиях. Хотя некоторые из этих признаков могли не проявляться в исходной популяции, гены, их кодирующие, уже в ней присутствовали. (Пока оставим в стороне мутации, так как многие новые разновидности могут возникать и без них.)
Такой тип изменчивости возможен только при наличии исходного хранилища информации, из которого можно вести отбор. Естественный отбор объясняет выживание наиболее приспособленных, но не отвечает на вопрос, откуда берутся новые, ранее не существовавшие приспособления. Менделевская изменчивость ничего не говорит о происхождении самой информации в популяции. Поэтому для демонстрации эволюции «вверх» требуются изменения иного рода, поскольку добавления новых полезных «предложений» не происходит. Это верно даже при возникновении новых видов. Если определять вид как репродуктивно изолированную единицу, то новые виды могут появляться без каких-либо «восходящих» изменений, то есть без добавления новой информации для новых функций. Например, мутация, вызывающая дефект, может создать генетический барьер или такие физические различия (как между догом и чихуахуа), которые сделают естественное скрещивание невозможным.
Удивительно, но некоторые креационисты до сих пор придерживаются взгляда, что новые виды никогда не появлялись. Однако полевые наблюдения часто демонстрируют «ряды» организмов, явно указывающие на то, что два репродуктивно изолированных вида произошли от общего предка. Те же креационисты, вероятно, согласятся, что волки, лисы, койоты и домашние собаки произошли от одного предка. Зачем же тогда отрицать возможность видообразования в настоящее время? В этом нет необходимости ни с библейской, ни с научной точки зрения. Более того, отрицание очевидных фактов лишь дает повод для критики.
Когда мы видим новые породы собак, мы наблюдаем не создание новой информации, а перераспределение и истощение уже имевшейся в исходной популяции. Попытка вывести чихуахуа из популяции догов обречена на провал — необходимые «генетические предложения» были утеряны в процессе специализированной селекции.
Таким образом, то, что кажется впечатляющим примером изменчивости, при ближайшем рассмотрении оказывается движением «вниз» — потерей информации и сокращением генетического потенциала. Экстраполяция такого процесса в будущее приведет не к эволюции, а к вырождению и вымиранию.
2. Полиплоидия
В этом случае также не возникает новых информационных предложений. Полиплоидия — это просто умножение («ксерокопирование») уже существующего генома.
3. Гибридизация
Здесь мы снова видим лишь смешивание двух уже существующих наборов информации, а не создание новой.
4. Мутации
Поскольку мутации по своей сути являются случайными нарушениями, большинство из них вредны, летальны или нейтральны.
Обратите внимание: Профессор Воробьев и дезинформация о вакцине «Спутник v».
Случайные ошибки в сложном генетическом коде вносят «шум» и хаос, а не новое осмысленное содержание или функцию. Однако в редких случаях разрушительное изменение может дать ситуативное преимущество. Например, мутация, укорачивающая ноги овцы, не даст ей перепрыгнуть через забор. Или потеря крыльев у жука на ветреном острове спасет его от сноса в море. Слепые пещерные рыбы или устойчивость к антибиотикам — это примеры «полезных» мутаций, которые не увеличивают функциональную сложность, а часто связаны с утратой функции. Случайные изменения не могут систематически создавать новую сложность.Здесь может возникнуть возражение о злоупотреблении терминами «полезный» или «функциональный». Критики скажут, что если изменение повышает выживаемость, то оно по определению полезно. Но такой подход сводит цель живых систем исключительно к выживанию, игнорируя тот факт, что они и их подсистемы выполняют специфические, замысловатые функции. Другими словами, они несут телеономическую информацию (информацию о цели или функции). Это ключевое отличие живого от неживого (за исключением машин, созданных человеком). Функции живых систем не всегда напрямую связаны с выживанием, но они выполняются эффективно. Эволюционист, конечно, может допустить, что когда-то в прошлом любая функция давала преимущество, но такие допущения непроверяемы. Поэтому законно говорить о генетической информации в телеономическом смысле, независимо от ее сиюминутной пользы для выживания.
Современные генофонды несут огромное количество информации, кодирующей сложные функции, которых не было в гипотетической «первой клетке». Чтобы обосновать эволюцию от простого к сложному, необходимо показать примеры мутаций, которые добавляют новые «информационные предложения» для новых функций.
На сегодняшний день нам не известна ни одна мутация, приводящая к такому возрастанию функциональной сложности. Соображения теории вероятности также делают такое событие крайне маловероятным, даже в масштабах миллиардов лет.
Для иллюстрации рассмотрим молекулу гемоглобина. Существует мутация (замена одной «буквы» в генетическом коде), которая в гомозиготном состоянии вызывает серповидно-клеточную анемию — тяжелое заболевание. Однако если мутация унаследована только от одного родителя (гетерозиготное состояние), она дает некоторую устойчивость к малярии. Эволюционисты часто приводят этот пример как «полезную мутацию». Но что мы видим на самом деле? Функционально эффективная молекула гемоглобина превратилась в поврежденную. Мы не наблюдаем улучшения механизма переноса кислорода или создания нового. Это не движение «вверх».
Простая аналогия: секретный агент ежедневно отправляет телеграмму: «Сегодня враг не готовится к атаке». Ошибка, в результате которой теряется частица «не», в большинстве случаев приведет к катастрофе. Лишь в исключительной ситуации она может случайно оказаться «полезной». Но такое изменение никак не превратит короткую телеграмму в объемную энциклопедию.
Крайне малое число «полезных» мутаций, наблюдаемых в природе, — это не тот тип изменений, который требуется для эволюции. Мы не видим добавления новых осмысленных информационных предложений. Интересно, что некоторые креационисты, отрицая любую пользу мутаций, также попадают в ловушку, если определять пользу лишь как повышение выживаемости в конкретных условиях.
Информация связана с упорядоченностью. Все наши наблюдения и законы термодинамики (рост энтропии) говорят о том, что в спонтанных, неуправляемых процессах упорядоченность уменьшается. То же самое относится и к информации. Физик или инженер связи не удивится, узнав, что биологические процессы не сопровождаются увеличением полезной, функциональной информации. Конечный результат любого процесса наследственной изменчивости — это сохранение или потеря генетической информации.
Это указывает на то, что исходный объем информации был создан сверхъестественным образом. Такой взгляд полностью согласуется с библейской картиной мира, сотворенного «хорошо весьма», чей упадок начался после грехопадения. Вот почему существуют пределы изменчивости, и креационисту не нужно беспокоиться о том, сколько новых «видов» появится в будущем — изначальный запас функциональной информации был конечен, и естественные процессы не могут его увеличить.
Заметим, что поскольку организмам предстояло расселиться по Земле и заполнить различные экологические ниши в условиях меняющегося мира, они изначально должны были обладать значительным потенциалом генетической изменчивости. Без такой заложенной пластичности многие популяции не выжили бы. Таким образом, сама концепция биологической изменчивости предсказывается библейской моделью, а не навязывается ей постфактум.
Природа и границы сотворенного рода
Писание подразумевает, что изначально сотворенная информация была распределена не в одном «супер-виде», а в нескольких отдельных генофондах — сотворенных родах. Каждая такая группа, размножающаяся половым путем, состояла как минимум из двух особей. Следовательно:
- Каждый первоначальный род начинал с уже встроенного запаса генетической информации, который стал основой для всей последующей полезной изменчивости.
- Каждый род, вероятно, был генетически и репродуктивно изолирован от других, но особи внутри рода свободно скрещивались. Таким образом, первоначальные сотворенные роды могли соответствовать современному понятию «вид».[4] Как мы видели на примере собак, такие «виды» могли делиться на подгруппы, которые расходились, теряли способность к скрещиванию и сами становились видами. Чем больше был исходный генетический потенциал, тем легче шло такое разделение. Однако каждый «раскол» уменьшал потенциал для дальнейших изменений. Все потомки одного первоначального рода могут в итоге быть классифицированы как одно крупное семейство.
Рассмотрим гипотетический сотворенный род А — биологический «вид» с огромным генетическим потенциалом (см. Рис. 1). Заметим, что род А мог сохраняться, как и некоторые из его дочерних популяций. Отщепление новых групп не обязательно ведет к вымиранию родительской формы. В случае человека первоначальная группа не дивергировала до уровня образования новых видов.
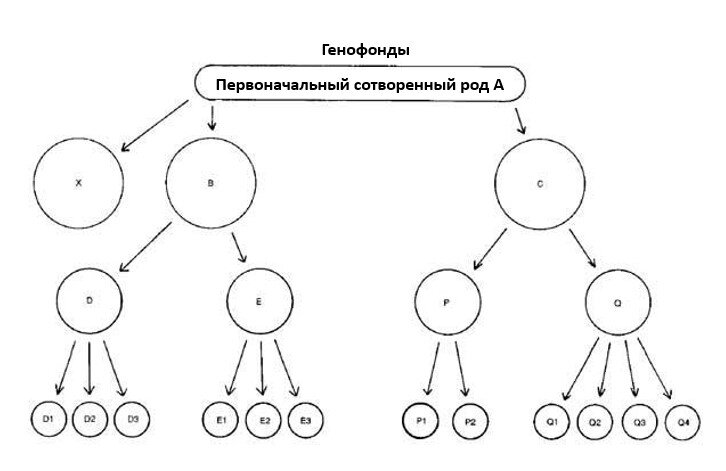 Рисунок 1. Схематическое изображение дивергенции дочерних популяций от первоначального сотворенного рода.
Рисунок 1. Схематическое изображение дивергенции дочерних популяций от первоначального сотворенного рода.В такой модели D1, D2, D3, E1, E2, E3 и т.д. могут быть разными репродуктивно изолированными видами. Однако вся функциональная генетическая информация, содержащаяся в них, уже присутствовала в исходном роде А (плюс, возможно, накопленные мутационные дефекты).
Предположим, что род А и некоторые промежуточные популяции (B, C, D, E) вымерли. Если какая-то часть информации из А была уникальна и не передалась потомкам, то она утрачена навсегда. Таким образом, несмотря на появление «новых видов», мы наблюдаем сохранение, перераспределение и потерю информации, но не добавление новой (полезной) информации. Это не тот процесс, который может объяснить эволюцию от простейших к человеку.
Таксономические категории выше вида (род, семейство и т.д.) — это в значительной степени произвольные группировки, основанные на морфологическом сходстве. Вполне возможно, что потомки одного сотворенного рода (например, D1, D2, D3) сегодня будут отнесены к одному роду, а потомки другого (E1, E2, E3) — к другому. Группы B и C могли разойтись настолько, что их потомки окажутся в разных семействах. Это создает проблему для креациониста, который пытается провести четкие границы между современными представителями разных бараминов.
Организмы могут быть отнесены к одному семейству по морфологическому сходству, но при этом происходить от двух разных сотворенных родов. Это предостерегает против использования только морфологии для классификации и указывает на ошибочность жестких заявлений типа: «в этом случае барамин — это семейство, а в том — род».
Простого решения для определения генетических взаимосвязей всех групп не существует, и, вероятно, мы никогда не узнаем их с абсолютной точностью. Эксперименты по скрещиванию или анализу ДНК могут указывать на общее происхождение, но отсутствие совместимости не доказывает обратного (генетические барьеры могли возникнуть из-за мутаций). Тем не менее, современные методы анализа последовательностей ДНК открывают новые возможности для таких исследований (хотя вопрос финансирования в доминирующей эволюционной парадигме остается открытым).
Итак, что ответить эволюционисту, который спрашивает: «Дайте определение сотворенному роду и скажите, как его опознать?» Предлагается следующее определение:
- «Группы организмов принадлежат к одному сотворенному роду, если они произошли от одного предкового генофонда».
Это определение аксиоматично и, подобно свернувшемуся в клубок броненосцу, является замкнутым. Оно переводит внимание с вопроса «Что такое барамин?» на более важный: «Какие современные популяции связаны общим происхождением от одного исходного генофонда?» Заметим, что это понятие общего происхождения радикально отличается от эволюционного. Креационист, глядя в прошлое, видит расширяющийся генофонд (больше информации в исходном роде). Эволюционист видит сужающийся, примитивный генофонд.
Как и во всех таксономических вопросах, здравый смысл будет играть ключевую роль. Писание, данные палеонтологии и здравый смысл, вместе взятые, не позволят креационистам «сваливать все в кучу». Например, разумно предположить, что крокодилы и аллигаторы произошли от общего предкового генофонда, но вряд ли этот же генофонд содержал гены, необходимые для страуса.
Ссылки и примечания
1. Lewin, R., Evolutionary theory under fire, Science 210(4472):883–887, 1980.
2. Lewin, ref. 1, p. 884.
3. Транспозоны, или «прыгающие гены», которые переносят генетическую информацию внутри генома, в данной дискуссии не рассматриваются, так как исследования в этой области еще не достигли зрелости, чтобы стать «новой надеждой» эволюции. Стоит отметить, что эти процессы, как и мутации, по-видимому, являются случайными и ненаправленными и связаны с перемещением уже существующей информации.
4. Первоначальный род мог быть представлен двумя или более географически изолированными, но потенциально скрещивающимися популяциями. При строгом определении вида их можно было бы считать одним «сотворенным» видом. Некоторые креационисты называют такие изоляты «морфотипами» внутри барамина.
Еще по теме здесь: Новости науки и техники.
Источник: Изменчивость, информация, и сотворенный род.